ДЕЗОКСИРИБОНУКЛЕИ́НОВЫЕ КИСЛО́ТЫ
-

Рубрика: Биология
-

-

Скопировать библиографическую ссылку:
ДЕЗОКСИРИБОНУКЛЕИ́НОВЫЕ КИСЛО́ТЫ (ДНК), нуклеиновые кислоты, содержащие в качестве углеводного компонента дезоксирибозу. ДНК – осн. компонент хромосом всех живых организмов, вещество, из которого построены геномы всех про- и эукариот, а также внехромосомные наследств. элементы (плазмиды) и геномы мн. вирусов. В клетках прокариот ДНК организована в виде компактного образования – нуклеоида. У эукариот она содержится в ядрах клеток и в органеллах – митохондриях и хлоропластах. В нуклеотидной последовательности ДНК записана (кодирована) генетич. информация о всех признаках вида и особенностях индивидуума. Все осн. генетич. процессы – репликация, транскрипция и рекомбинация связаны с функционированием молекулы ДНК.
Впервые ДНК в виде комплексов с белками (дезоксирибонуклеопротеидов) была открыта в 1868 И. Ф. Мишером в ядрах клеток гноя и сперме рыб. Долгое время считалось, что ДНК содержится только в клетках животных, и лишь к сер. 1930-х гг. было доказано (А. Н. Белозерский), что ДНК – непременный компонент каждой живой клетки. В 1944 амер. микробиолог О. Эйвери с сотрудниками показали, что с помощью ДНК, тщательно очищенной от всех остальных клеточных компонентов, наследуемый биологич. признак может быть перенесён из одной клетки в другую. Тем самым была определена биологич. функция ДНК как вещества наследственности.
В кон. 19 – нач. 20 вв. было установлено, что ДНК представляют собой полимерные молекулы, мономерными составляющими которых служат дезоксирибонуклеотиды, содержащие остатки дезоксирибозы, фосфорной кислоты и одно из четырёх азотистых оснований: пуриновых – гуанина (G) и аденина (А) и пиримидиновых – цитозина (C) и тимина (Т). В кон. 1940-х – нач. 1950-х гг. в лаборатории А. Тодда было доказано, что единственным типом межнуклеотидной связи в полимерных цепях ДНК является 3'–5'-фосфодиэфирная связь. В это же время Э. Чаргафф с сотрудниками выяснили осн. закономерности нуклеотидного состава ДНК (правила Чаргаффа), наиболее важная из которых – равенство содержания остатков аденина и тимина (А = Т), а также гуанина и цитозина (G = C).
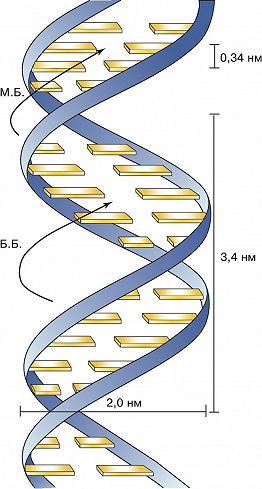
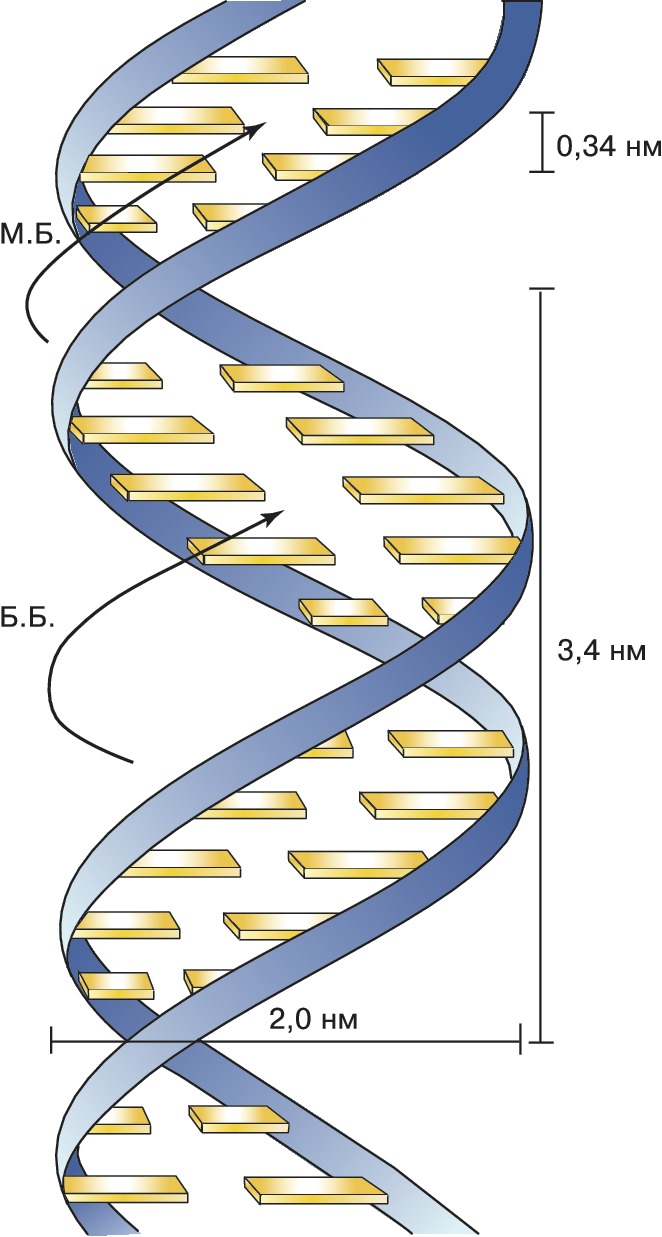
Основываясь на этих данных, в 1953 Дж. Уотсон и Ф. Крик расшифровали рентгенограммы кристаллов ДНК, полученные в лабораториях Р. Франклин и М. Уилкинса, и сделали одно из выдающихся открытий совр. естествознания. Они установили, что молекула ДНК представляет собой регулярную спираль, состоящую из двух полинуклеотидных цепей (двойная спираль). Диаметр спирали постоянен на протяжении всей её длины и равен примерно 2 нм. Длина витка спирали составляет 3,4 нм. На один виток в одной цепи приходится примерно 10 нуклеотидных остатков, т. е. межнуклеотидное расстояние вдоль оси спирали равно 0,34 нм. Азотистые основания в двойной спирали ДНК лежат в одной плоскости, которая практически перпендикулярна её гл. оси. При этом основания, принадлежащие разным цепям и находящиеся напротив друг друга, образуют комплементарные пары, стабилизированные водородными связями таким образом, что аденин всегда спарен только с тимином, а гуанин – с цитозином (пары G – C связаны между собой тремя водородными связями, а пары А – Т лишь двумя). Для стабилизации структуры двойной спирали ДНК важное значение имеют также взаимодействия между плоскостями соседних оснований, принадлежащих одной и той же цепи (т. н. стэкинг-взаимодействия, от англ. stack – стог, складывать в стог, располагать один над другим).
Из модели двойной спирали Уотсона – Крика прямо вытекает принцип самовоспроизведения (удвоения, репликации) молекулы ДНК (а следовательно, и любого генетич. материала): если две комплементарные цепи ДНК разделить, а затем на каждой, как на матрице, построить новые, строго комплементарные им цепи, то две дочерние двуспиральные молекулы будут идентичны материнской. Открытие этого принципа позволило на молекулярном уровне объяснить явление наследственности и положило начало молекулярной биологии. Принцип комплементарного спаривания оснований нуклеиновых кислот лежит в основе всех процессов передачи генетич. информации в клетке.
В двойной спирали ДНК сахарофосфатный остов полинуклеотидных цепей обращён наружу, а на поверхности спирали можно выделить две бороздки: большую – шириной 2,2 нм и малую – шириной 1,2 нм. Двойная спираль ДНК, описанная Дж. Уотсоном и Ф. Криком, – правозакрученная, а полинуклеотидные цепи в ней антипараллельны, т. е. направлены в противоположные стороны, так что 3'-конец одной цепи располагается напротив 5'-конца другой. Она была названа В-формой ДНК.
Оказалось, однако, что двойная спираль ДНК характеризуется существенным полиморфизмом и при изменении внешних условий может принимать пространственную структуру (конформацию), отличную от уотсон-криковской В-формы. Так, при понижении влажности в препарате или, напр., при добавлении спирта к водному раствору ДНК она переходит в т. н. А-форму, отличающуюся от В-формы шириной и глубиной бороздок, увеличением диаметра спирали, смещением пар оснований к периферии спирали и их заметным наклоном по отношению к оси спирали, а внутри неё образуется полость диаметром 0,4 нм. В основе этих структурных превращений лежит изменение конформации остатка дезоксирибозы, что, в свою очередь, ведёт к изменению расстояния между фосфатными группами соседних нуклеотидных остатков одной цепи. При высокой концентрации солей участки двойных спиралей ДНК с чередующимися нуклеотидными последовательностями типа многократно повторяющегося гуанозин-цитозинового динуклеотида (GC) из правозакрученной формы переходят в левозакрученную. У этой формы ДНК линия, соединяющая фосфатные группы, через каждые две пары имеет излом и принимает зигзагообразный вид. Такая конформация ДНК называется Z-формой (от англ. zigzag). Хотя полиморфизм ДНК может играть существенную роль в регуляции активности генов, прямых данных о наличии у двойной спирали ДНК in vivo иных конформаций, кроме В-формы, пока нет.
Важным свойством двойных спиралей ДНК является их микрогетерогенность, обнаруживаемая рентгеноструктурным анализом высокого разрешения. Она обусловлена тонкими различиями в конформации нуклеотидных остатков, появление которых зависит от последовательности расположения нуклеотидов в цепи, и проявляется в образовании характерных изгибов и изломов. Такие особенности структуры молекулы ДНК, несомненно, должны быть связаны с её функционированием.
При наличии в молекуле ДНК повторяющихся последовательностей (палиндромов) могут формироваться пары не только между основаниями противоположных цепей, но и в пределах одной цепи, что создаёт возможность образования связанных водородными связями своеобразных шпилек с петлями.
При повышении темп-ры или рН растворов ДНК, в присутствии ряда органич. веществ и др. соединений происходит денатурация ДНК – разрыв водородных связей между парами оснований и разрушение регулярной структуры двойной спирали, которое завершается полным разделением цепей. Благодаря кооперативному характеру внутримолекулярных взаимодействий, стабилизирующих двойную спираль, этот процесс напоминает фазовый переход и поэтому называется плавлением ДНК. В условиях, оптимальных для образования двойной спирали, отд. комплементарные цепи ДНК способны реассоциировать с восстановлением исходной двуспиральной структуры (ренатурация ДНК). Это свойство лежит в основе метода молекулярной гибридизации нуклеиновых кислот, который позволяет выявлять степень сходства нуклеотидных последовательностей молекул ДНК или ДНК и РНК, особенности их организации, в т. ч. наличие и число повторов (см. Нуклеотидные последовательности).
Последовательность чередования нуклеотидных остатков в ДНК (первичная структура) у разных организмов строго индивидуальна и служит важнейшей характеристикой, отличающей одну молекулу ДНК от другой и соответственно один ген или один регуляторный генетич. элемент от другого. Размеры молекул ДНК варьируют от нескольких тысяч пар нуклеотидов (т. п. н.) у плазмид и некоторых вирусов до сотен т. п. н. у высших организмов. Содержание ДНК в разных организмах также различно и по числу образующих её нуклеотидов составляет от 5·106 у бактерий до 2·1011 пар нуклеотидов (п. н.) у высших растений (в расчёте на гаплоидный геном). Эти гигантские молекулы чрезвычайно компактно упакованы в клетках или вирусах. В прокариотич. нуклеотиде такая укладка поддерживается небольшим количеством спец. белков и, вероятно, рибонуклеиновыми кислотами (РНК). Описано неск. уровней упаковки эукариотической ДНК с помощью универсального набора гистонов и некоторых негистоновых белков, приводящих к образованию осн. компонента хромосомы – хроматина. Напр., длина ДНК самой большой хромосомы человека равна 8 см, но в хромосоме (в состоянии митоза) она не превышает 5 мкм.
В ядрах эукариот (за исключением гамет) ДНК представлена двумя копиями. Каждая про- и эукариотическая хромосома содержит только одну молекулу двуспиральной ДНК. Геном подавляющего большинства вирусов также представлен двуспиральной ДНК, и лишь некоторые фаги в качестве геномной содержат однотяжевую кольцевую или линейную молекулу ДНК.
В кольцо замкнуты молекулы двунитевых ДНК прокариотич. хромосомы, плазмид и мн. вирусов, ДНК митохондрий и хлоропластов. При этом если цепь ковалентно-непрерывна (т. е. все фосфодиэфирные связи замкнуты), то циклич. ДНК могут находиться в сверхспирализованной форме, когда нити двойной спирали многократно зацеплены друг с другом. В клетке сверхвитки создаются и разрушаются ферментами топоизомеразами. Циклическая сверхспирализованная ДНК обладает определённым запасом энергии по сравнению с её линейной формой, поэтому образование сверхвитков важно для функционирования ДНК (напр., позволяет разрешать топологич. трудности, возникающие при репликации). Кроме того, благодаря наличию сверхвитков могут образовываться необычные структуры в её макромолекуле: крестообразные структуры (в палиндромах), Z-форма, тринитевые участки, или т. н. Н-форма (в гомопурин-гомопиримидиновых блоках).
Биосинтез ДНК (репликация) осуществляется путём матричного синтеза при участии ферментов ДНК-полимераз совместно с большой группой вспомогат. белков и находится под контролем спец. регуляторных систем клетки. In vitro любой участок ДНК может быть амплифицирован с помощью полимеразной цепной реакции. В ходе репликации in vivo, а также после её окончания происходит метилирование небольшого числа определённых остатков цитозина с образованием 5-метилцитозина, представляющее собой специфич. процесс модификации ДНК, непосредственно связанный с её последующим функционированием. Метилирование и деметилирование ДНК играют важную роль в процессах эмбрио- и гаметогенеза.
В ходе жизнедеятельности организмов их ДНК под влиянием внешних факторов может подвергаться разл. повреждающим воздействиям, сопровождающимся нарушением структуры азотистых оснований. В ходе эволюции клетки выработали защитные механизмы, обеспечивающие восстановление исходной структуры – репарацию ДНК.
В клетке ДНК расщепляется специфич. ферментами – дезоксирибонуклеазами. Среди них наиболее известны эндонуклеазы рестрикции, защищающие клетку от чужеродной ДНК и широко применяемые в генетич. инженерии.
В нач. 1970-х гг. Ф. Сенгером и др. были разработаны эффективные методы определения последовательности нуклеотидов в молекулах ДНК (см. Секвенирование). В кон. 20 в. на основе этих методов создана мощная автоматизир. технология секвенирования ДНК, с помощью которой определена первичная структура ДНК полных геномов мн. вирусов, митохондрий, хлоропластов, бактерий, растений и животных. К 2004 было завершено определение нуклеотидной последовательности практически всего генома человека (более трёх млрд. п. н.). Эти работы стимулировали развитие биоинформатики и положили начало новому разделу молекулярной генетики – геномике.
Информация о нуклеотидных последовательностях ДНК широко используется при создании рекомбинантных ДНК – молекул с заданными свойствами, включающих структурные элементы ДНК разных организмов (см. Генетическая инженерия), а также при конструировании новых белков (см. Белковая инженерия). Знание первичной структуры ДНК важно при анализе наследств. и онкологич. заболеваний, идентификации личности (см. ДНК-типирование), при амплификации и выделении определённых генов, регуляторных элементов и др. функционально важных участков ДНК.
-
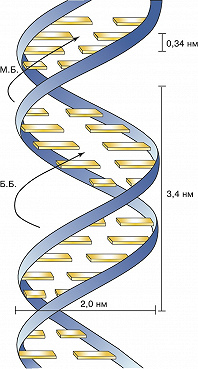 Рис. 1. Модель двойной спирали ДНК Уотсона – Крика (М. Б. – малая бороздка; Б. Б. – большая бороздка).
Рис. 1. Модель двойной спирали ДНК Уотсона – Крика (М. Б. – малая бороздка; Б. Б. – большая бороздка). -
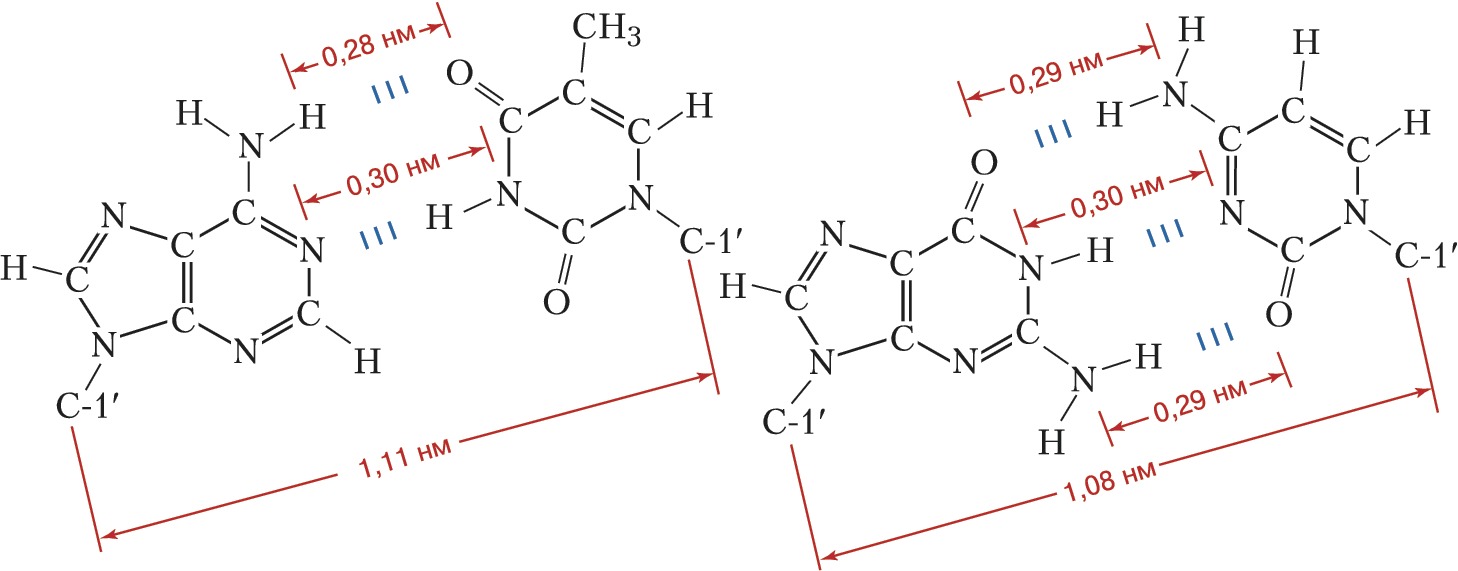
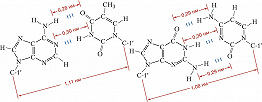
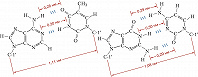 Рис. 2. Комплементарные уотсон-криковские пары в двуспиральной молекуле ДНК. Слева – пара аденин – тимин; справа – пара гуанин – цитозин. Приведены расстояния между атомами, связанными специфическими водородными связями, а также расстояния между С-1'атомами остатков дезоксирибозы.
Рис. 2. Комплементарные уотсон-криковские пары в двуспиральной молекуле ДНК. Слева – пара аденин – тимин; справа – пара гуанин – цитозин. Приведены расстояния между атомами, связанными специфическими водородными связями, а также расстояния между С-1'атомами остатков дезоксирибозы.