ДНК-ТИПИ́РОВАНИЕ
-

Рубрика: Биология
-

-

Скопировать библиографическую ссылку:
ДНК-ТИПИ́РОВАНИЕ (молекулярно-генетическая индивидуализация организмов, ДНК-индивидуализация), способ исследования генетич. материала, направленный на выявление и оценку индивидуальных генетич. особенностей биологич. объекта. Цель такого исследования – установление сходства (или различий) разных организмов для определения степени их родства.
Метод ДНК-т. основан на существовании различий в структуре ДНК у разных индивидуумов. Это касается т. н. гипервариабельных участков, обладающих структурным полиморфизмом (имеют неск. аллельных форм). В их формировании гл. роль играют относительно короткие нуклеотидные последовательности, получившие назв. минисателлитные и микросателлитные ДНК, состоящие соответственно из 15–70 и 2–5 пар нуклеотидов. Эти последовательности рассеяны по геному в виде блоков (локусов) и имеют тандемную организацию (многократно следуют друг за другом). Число тандемных повторов (и, следовательно, длина самих локусов) варьирует в пределах от 2–4 до нескольких тысяч. Наличие повторяющихся элементов в таких гомологичных сателлитных блоках и обусловливает структурный полиморфизм этих локусов, проявляющийся, в частности, как феномен полиморфизма длины рестриктазных фрагментов (ПДРФ). Такие фрагменты образуются после ферментативного расщепления ДНК рестриктазами (внутри повторов расщепления не происходит из-за особенностей их нуклеотидного состава). В первоначальных вариантах ДНК-т. фрагменты ДНК разделяли электрофорезом в агарозном геле, а затем получали отпечаток этого геля на мембранном фильтре. Последний инкубировали в растворе, содержащем специально подобранные зонды – фрагменты ДНК, меченные радиоизотопом. Содержащие комплементарные зонду участки исследуемой ДНК связывались (гибридизовались) с ним и выявлялись с помощью радиоавтографии. Т. о. на радиочувствительной плёнке в виде набора полос идентифицировались сразу все микро- и минисателлиты геномной ДНК, гомологичные используемому зонду. По аналогии с анализом дактилоскопич. оттиска (рисунком папиллярных линий) появился термин «генетическая, или геномная, дактилоскопия». Каждый индивидуум имеет свой геномный «отпечаток», который характеризуется определённым числом полос (до нескольких десятков), их расположением на дорожке и интенсивностью почернения каждой из полос. Чем больше родство, тем больше совпадений. Вся картина обнаруживает даже более высокую индивидуальную специфичность, чем папиллярные узоры, и поэтому может служить «генетич. удостоверением» личности. У кровных родственников число совпадающих полос заметно больше, у близнецов они идентичны.
Разработка метода (кон. 1980-х гг.) полимеразной цепной реакции (ПЦР) позволила размножать (амплифицировать) любую последовательность ДНК, получать десятки и сотни миллионов её копий и использовать для молекулярно-генетич. анализа исчезающе малые количества биологич. материала. Появилась возможность сравнивать длину отд. целых локусов минисателлитных ДНК, не подвергая их предварительному ферментативному расщеплению, т. е. осуществлять анализ полиморфизма длины амплифицированных фрагментов (ПДАФ). После разделения продуктов ПЦР электрофорезом их положение в геле выявляют с помощью спец. красителей, флуоресцентных меток и др. Разработаны также т. н. мультиплексные амплификационные системы, которые позволяют одновременно анализировать ПДАФ сразу нескольких локусов (их может быть более десяти). В этом случае амплификационный профиль ДНК, создаваемый уже суммой локусов, оказывается чрезвычайно полиморфным (является комбинацией нескольких независимых полиморфных элементов), но обеспечивает очень высокую достоверность анализа.
Вариабельность в структуре полиморфных генов мини- и микросателлитных ДНК может быть обусловлена также точечными нуклеотидными заменами. Такие локусы различаются только по нуклеотидному составу. В этом случае определяют первичную структуру амплифицированных фрагментов (см. Секвенирование). Наиболее разработанным методом ДНК-т., основанным на секвенировании фрагментов, является анализ митохондриальной ДНК (мтДНК), которая характеризуется высоким уровнем полиморфизма, наличием большого числа копий, отсутствием рекомбинации и материнским характером наследования (зародыш получает митохондрии только из яйцеклетки). Всё это позволило широко использовать мтДНК в популяционных и филогенетич. исследованиях, а в некоторых случаях сделало её единственно возможным инструментом для ДНК-т.; напр., когда ДНК, содержащаяся в образце, сильно деградирована, а хромосомная ДНК не может быть амплифицирована. Наследование по материнской линии и отсутствие рекомбинации позволяют использовать мтДНК в качестве т. н. «трассирующего» родословного генетич. маркера. Это особенно важно при установлении родства в тех случаях, когда генетич. дистанция, разделяющая родственников, оказывается больше, чем одно поколение. Ярким примером использования мтДНК для ДНК-т. явилась работа рос., брит. и амер. учёных по идентификации останков рос. имп. семьи в 1992–98. Позднее анализ мтДНК был многократно использован для индивидуализации костных и мумифицир. останков, возраст которых исчисляется десятками, сотнями и даже десятками тысяч лет.
Методом ДНК-т. пользуются в криминалистике для идентификации личности и установления родств. отношений, а также в медико-биологич. анализе. В генетике и селекции с.-х. животных и растений этот метод используют для паспортизации и отбора чистопородного потомства животных, типирования сортов растений, определения межвидовых и межсортовых различий, а в практич. бактериологии и эпидемиологии – для идентификации бактериальных штаммов. ДНК-т. можно использовать при изучении структурно-функциональных особенностей генетич. аппарата и явлений геномной нестабильности при нормальном функционировании клеток и патогенезе, в популяционной и эволюционной генетике.
-
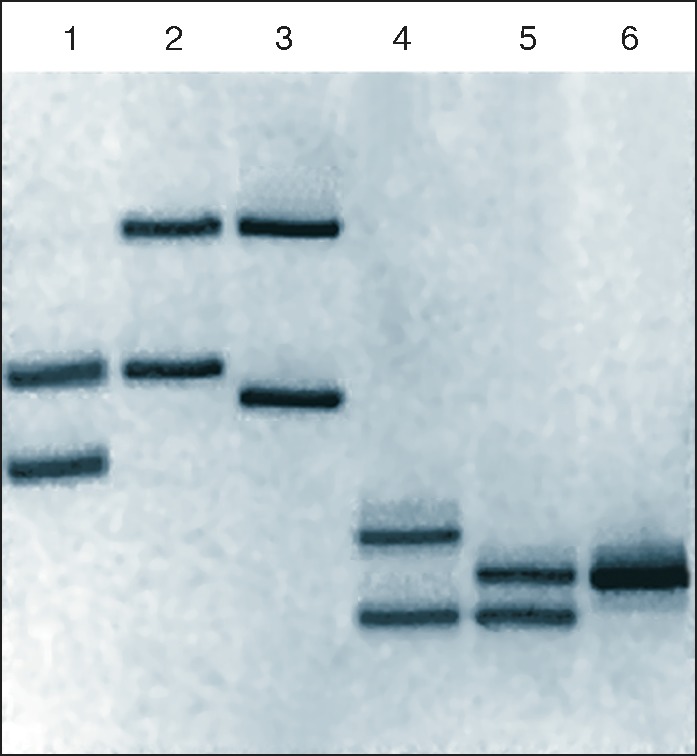
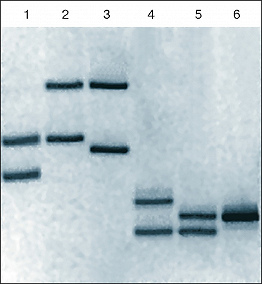
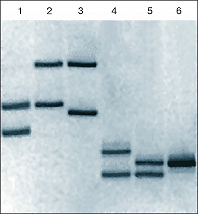 Электрофореграмма двух амплифицированных фрагментов ДНК: аллели локусов 3’A’poB (дорожки 1–3) и D1S80 (дорожки 4–6) у семейной группы мать (дорожки 1, 4), ребёнок (дорожки 2, 5), отец (дорожки 3, 6). Демонстрирует наличие одинаковых фрагментов генетического материала родителей у ребёнка и полное их отсутствие у матери и отца – неродственных индивидуумов.
Электрофореграмма двух амплифицированных фрагментов ДНК: аллели локусов 3’A’poB (дорожки 1–3) и D1S80 (дорожки 4–6) у семейной группы мать (дорожки 1, 4), ребёнок (дорожки 2, 5), отец (дорожки 3, 6). Демонстрирует наличие одинаковых фрагментов генетического материала родителей у ребёнка и полное их отсутствие у матери и отца – неродственных индивидуумов.