ВИ́РУСЫ
-

Рубрика: Биология
-

-

Скопировать библиографическую ссылку:
ВИ́РУСЫ (от лат. virus – яд), мельчайшие биологич. объекты (обычно размером от сотых до десятых долей мкм). Важнейшая особенность В. – их неспособность к самостоятельному размножению, что связано с отсутствием у них механизмов для синтеза белков и производства энергии. Ранее считалось, что В. могут размножаться только в живых клетках (что обычно и имеет место), и это свойство входило в их науч. определение. Сейчас доказана принципиальная возможность размножения некоторых В. и в разрушенных клетках, но, несмотря на это, считается, что В. – облигатные (безусловные) внутриклеточные паразиты.
Строение вирусов
Вирусные частицы, или вирионы, могут иметь сферическую, нитевидную, палочковидную и более сложные формы. Одни В. (их иногда называют простыми) представлены только нуклеокапсидом – геномом В., заключённым в белковую оболочку (капсид), обычно построенную из множества молекул белка одного и того же или разных типов, уложенных по закономерностям кубической или спиральной симметрии. У других (сложных) В. нуклеокапсид окружён наружной оболочкой (мембраной), обычно состоящей из липидов с вкраплениями белков и гликопротеинов. Геном В. представлен либо молекулой ДНК (ДНК-содержащие В.), либо одной или несколькими молекулами РНК (РНК-содержащие В.), причём у разных В. эти молекулы могут быть однонитевыми или двунитевыми, линейными или кольцевыми. Размеры генома варьируют обычно от нескольких тысяч до 30 тыс. нуклеотидов у РНК-содержащих В. и до нескольких сотен тысяч нуклеотидов у ДНК-содержащих. Самые мелкие геномы имеют всего неск. генов, которые кодируют белок (белки) вирусной оболочки и некоторые ферменты, принимающие участие в синтезе вирусных нуклеиновых кислот. В более крупных геномах (особенно у ДНК-содержащих В.) закодирована дополнительная информация, не всегда обязательная для сохранения жизнеспособности В., но обеспечивающая их более эффективное и независимое от внутриклеточных условий размножение. Напр., в ДНК-геноме мимивируса, поражающего амёб, заключено ок. 1200 генов, т. е. больше, чем в ДНК некоторых паразитич. микроорганизмов. Существуют т. н. дефектные В. (напр., В. гепатита дельта), которых называют паразитами «вдвойне»; у них нарушены те или иные системы репродукции, поэтому для образования потомства им требуется не только внутриклеточная среда, но и помощь со стороны др. полноценного В. (в данном случае – В. гепатита B). Для некоторых парвовирусов в качестве помощника выступают аденовирусы. В. могут поражать все живые организмы. В. бактерий называют бактериофагами (фагами). Описано неск. тысяч В. Их относят к десяткам семейств, среди которых выделяют роды и виды.
Размножение вирусов
Размножение вирусов протекает в неск. стадий. Сначала В. прикрепляется к клетке, что обычно требует наличия химич. сродства между поверхностью вирусной частицы и тем или иным компонентом клеточной поверхности (рецептором). Это взаимодействие весьма специфично, что является одной из причин высокой избирательности взаимодействия В. с клеткой (напр., В. полиомиелита или В. иммунодефицита человека – ВИЧ в естеств. условиях инфекционны только для человека). Но есть В. и с широким спектром хозяев, заражающие, напр., млекопитающих и насекомых (так, В. клещевого энцефалита размножается в организме человека и клеща). В клетки растений В. может проникать только после механич. повреждения (напр., насекомыми) клеточной стенки, т. к. она служит для них непреодолимым барьером. После взаимодействия В. с рецептором его геном (в свободном виде или в комплексе с белками) проникает внутрь клетки, где начинается синтез вирусных нуклеиновых кислот и белков (структурных компонентов вириона, а также вирусных ферментов и др. молекул, способствующих эффективному размножению вируса благодаря их взаимодействию с вирусными или клеточными макромолекулами). Последовательность образования этих двух типов молекул у разных В. варьирует. Однако всегда синтез вирусных белков направляется вирусными нуклеиновыми кислотами и происходит на рибосомах клетки. Синтез же вирусных нуклеиновых кислот осуществляется при участии либо вирусных, либо клеточных ферментов, нередко и тех и других. Затем вирусные нуклеиновые кислоты и структурные белки объединяются с образованием дочерних вирионов, которые покидают клетку, разрушая её или сохраняя целостность. Клеточная плазматическая мембрана сохраняется, напр., если В. покидает клетку путём т. н. почкования: «незрелый» вирион сначала обволакивается клеточной мембраной, включающей некоторые из вирусных белков, затем «отшнуровывается», а целостность клеточной мембраны восстанавливается. Т. о., для построения оболочки В. могут использоваться клеточные липиды и углеводы. Инфекц. процесс, инициированный одним вирионом, может приводить к образованию до 103–105 дочерних вирусных частиц.
Выделяют неск. осн. типов взаимодействия В. и клетки. При продуктивной литической инфекции заражённая клетка после образования вирусного потомства гибнет. В случае хронической персистентной (стойкой) инфекции заражённая клетка продолжает жить и делиться, постоянно поддерживая размножение В., хотя её функции могут претерпевать некоторые изменения. Такая инфекция может продолжаться длительное время без заметных внешних проявлений, не вызывая признаков заболевания, хотя могут иметь место определённые изменения регуляции клеточных процессов. Особый тип взаимодействия с клеткой наблюдается у т. н. умеренных В., к числу которых относятся бактериофаг лямбда и ряд В. человека и животных (напр., ВИЧ). Их геном ковалентно встраивается (интегрируется) в клеточную хромосому, превращаясь в элемент клеточного генома, и теряет способность к автономной репликации. В таком состоянии В. называют провирусом, а бактериофаг – профагом. У умеренных РНК-содержащих вирусов (ретровирусы) геном предварительно переходит в ДНК-форму. Экспрессия вирусных генов происходит так же, как и у генов клетки, а кодируемые ими белки могут влиять на свойства клетки, напр. на регуляцию её деления. В некоторых случаях встраивание вирусного генома в геном клетки может приводить к почти полному угнетению экспрессии клеточных генов. При этом клетка сохраняет жизнеспособность, но её свойства могут существенно меняться (напр., может произойти её злокачественная трансформация). Патологич. изменения клетки могут быть также связаны с угнетением синтеза РНК и клеточных белков, нарушением мембранной проницаемости и др. процессами, заканчивающимися во мн. случаях смертью клетки (цитопатический эффект).
В клетке и в целом организме существуют механизмы, препятствующие размножению В. и развитию вирус-индуцированной патологии, в т. ч. врождённый (напр., индукция интерферонов) и приобретённый (напр., образование антител) иммунитет. В свою очередь, мн. В. располагают механизмами, противодействующими клеточной защите. Так, некоторые В. подавляют секрецию интерферона заражённой клеткой, облегчая тем самым заражение соседних клеток вирусным потомством. Исход инфекции и судьба заражённой клетки и организма зависят от баланса противоположно направленных активностей В. и клетки. Поражение клеток в заражённом организме ведёт к нарушению работы соответствующих органов и тканей, проявляющихся в виде вирусных заболеваний. Некоторые из них имеют характер массовых эпидемий и даже пандемий и могут сопровождаться значительной смертностью (в т. ч. СПИД, грипп, ящур и др.). Важнейшим средством профилактики вирусных заболеваний является специфическая вакцинация.
Генетическая изменчивость вирусов, их эволюция
Изменчивость В. объясняет их способность «уходить» от иммунного ответа заражённого им организма (напр., в случае В. гриппа) и легко образовывать варианты, резистентные к лекарственным препаратам. Результатом изменчивости может быть также приобретение способности вызывать заболевания человека В., ранее патогенными только для животных. Нередко обнаруживается явное родство между отд. белками (напр., ферментами, осуществляющими синтез нуклеиновых кислот) у В., которые, на первый взгляд, не имеют между собой ничего общего. В ряде случаев заметно сходство между вирусными и клеточными белками. Из этого следует, что в ходе эволюции происходил (и, по-видимому, происходит) обмен генетич. информацией как между разными В., так и между В. и клеточными организмами. Ярким примером явления трансдукции – способности В. переносить гены или их фрагменты от одного организма к другому – может служить бактерия дифтерийная палочка, которая начинает вырабатывать токсин только после заражения её определённым фагом. Такой перенос генов (его называют горизонтальным) пытаются использовать для генной терапии путём создания на основе В. искусств. конструкций – векторов, способных вводить в заражаемую клетку здоровые или корректирующие гены.
Особую категорию составляют т. н. эндогенные В. Их геном в виде двунитевой ДНК, соответствующей геному ретровирусов, постоянно находится в составе клеточной хромосомы и функционирует как набор клеточных генов. Эндогенные В. могут быть полноценными, и тогда их экспрессия приводит к образованию В., способного заражать др. клетки (напр., В. рака молочных желёз мышей). В большинстве случаев, однако, эндогенные В. имеют дефектный геном, в котором некоторые вирусные гены отсутствуют или повреждены. Генетич. материал разнообразных эндогенных В. составляет весьма значительную долю хромосомной ДНК животных, в т. ч. человека. Они обнаружены и у растений. Считается, что эндогенные В. возникли в результате заражения половых клеток и последующей интеграции вирусной и клеточной ДНК. Такое заражение в процессе эволюции могло происходить многократно, и многие эндогенные В. считаются весьма древними.
Ввиду огромного разнообразия В. построение их эволюционного «древа» весьма затруднено. Среди мн. предположений о происхождении В. наибольшего внимания заслуживают два: или они возникли из обособившихся («одичавших») элементов клеточного генома, или в какой-то форме существовали уже на доклеточной стадии биологич. эволюции.
В. сыграли и продолжают играть выдающуюся роль как удобные модельные объекты для изучения общих закономерностей молекулярной биологии. Именно при изучении В. были расшифрованы важнейшие закономерности синтеза белков и нуклеиновых кислот и регуляции этих процессов, сформулированы мн. понятия молекулярной биологии и молекулярной генетики. См. также ст. Вирусология и лит. при ней.
-
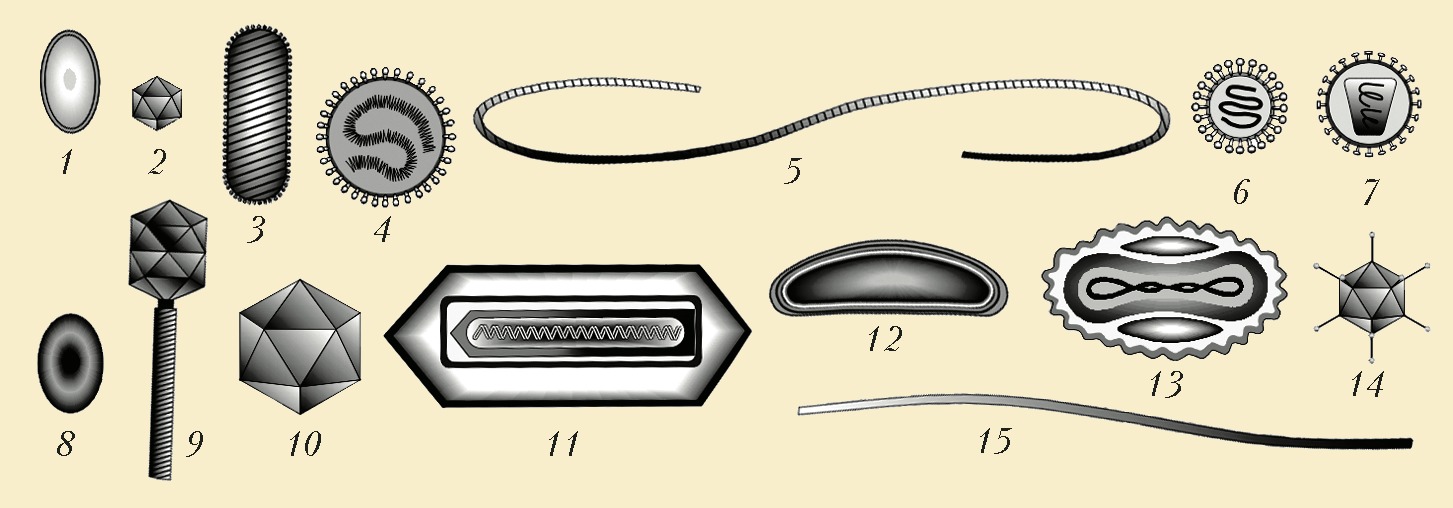
 Схемы строения представителей некоторых семейств вирусов. РНК-содержащие вирусы: 1 – гиповирус; 2 – реовирус; 3 – рабдовирус; 4 – парамиксовирус; 5 – клостеровирус; 6 – коронавирус; 7 – ретровирус. ДНК-содержащие вирусы: 8 – плазмавирус; 9 – миовирус; 10 – иридовирус; 11 – бакуловирус; 12 – асковирус; 13 – поксвирус; 14 – аденовирус; 15 – иновирус.
Схемы строения представителей некоторых семейств вирусов. РНК-содержащие вирусы: 1 – гиповирус; 2 – реовирус; 3 – рабдовирус; 4 – парамиксовирус; 5 – клостеровирус; 6 – коронавирус; 7 – ретровирус. ДНК-содержащие вирусы: 8 – плазмавирус; 9 – миовирус; 10 – иридовирус; 11 – бакуловирус; 12 – асковирус; 13 – поксвирус; 14 – аденовирус; 15 – иновирус.