БАКТЕРИОФА́ГИ
-

Рубрика: Биология
-

-

Скопировать библиографическую ссылку:
БАКТЕРИОФА́ГИ, фаги (от бактерии и греч. φάγος – пожиратель), вирусы бактерий. Открыты независимо англ. вирусологом Ф. Туортом (1915) и канад. бактериологом Ф. Д’Эррелем (1917). Являясь облигатными паразитами, Б. развиваются только в живых, активно метаболизирующих бактериях. Активная форма Б., т. н. вегетативный фаг, представляет собой его реплицирующийся и экспрессирующийся внутри клетки генетич. материал. Б. обнаружены во многих (но не во всех) исследовавшихся с этой целью видах бактерий. Как правило, каждый Б. развивается лишь в бактериях одного вида, хотя встречаются т. н. поливалентные Б., которые паразитируют в бактериях разных видов.
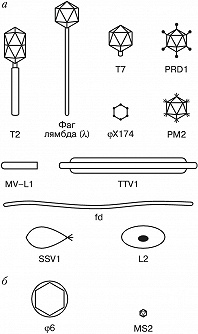
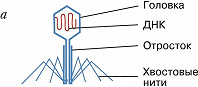
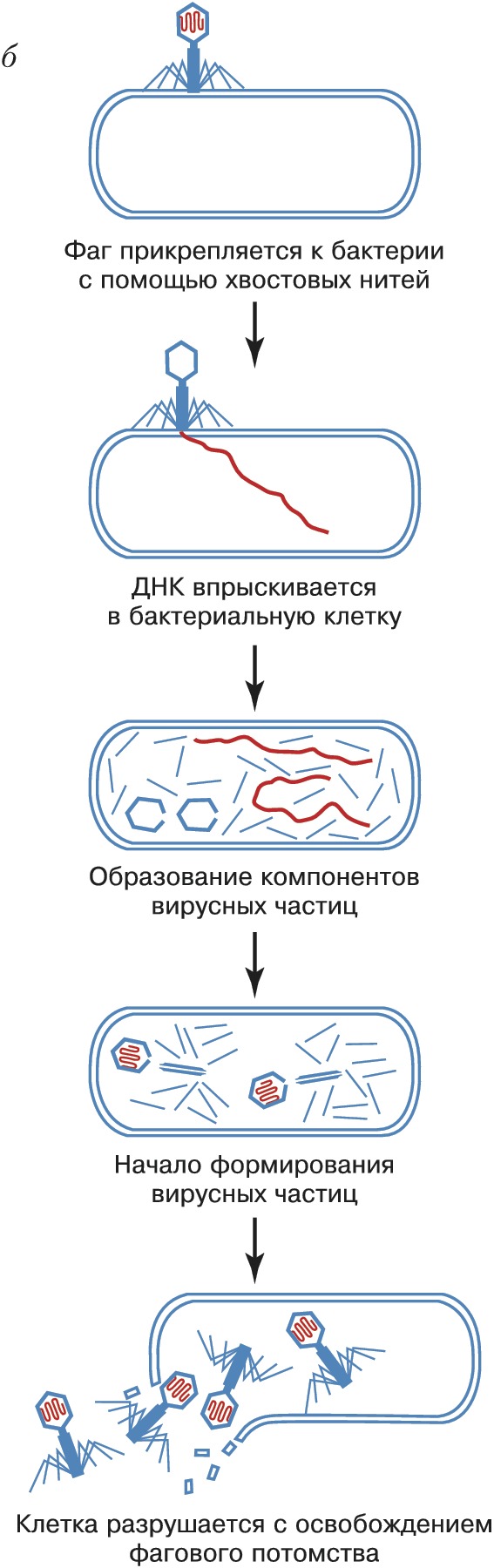
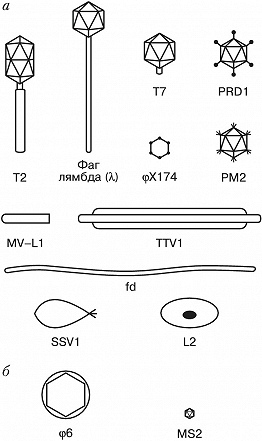
Фаговая частица состоит из молекулы нуклеиновой кислоты (геном Б.), заключённой в капсид – белковую оболочку, которая сохраняет геном вне клетки. Б. разнообразны по форме. Некоторые из них имеют вид многогранников с диаметром капсида 20–30 нм (φX174, F2, R17, MS2) или нитей длиной до 800 нм (fd, M13). Более сложно устроенные Б. (T-фаги, фаг лямбда и др.) имеют икосаэдрич. головку (диаметр ок. 100 нм) и отросток («хвост»), с помощью которого осуществляется контакт с клеткой.
Б. классифицируют, исходя из морфологич. особенностей частиц (форма головки, наличие хвоста, его размер и способность к сокращению) и сходства геномов и контролируемых ими белковых продуктов. Геномы разных Б. включают от неск. генов у мелких Б. до неск. сотен генов у гигантских вирулентных Б. У большинства Б. геном представлен двухцепочечной ДНК. Описаны также Б. с одноцепочечной ДНК (M13, S13, φ X174) или РНК (MS2, Qβ ). Молекулы нуклеиновых кислот могут иметь разные размеры и находиться в линейной, фрагментированной или кольцевой форме. У некоторых Б. (напр., фаг лямбда) геном имеет линейную форму, но после попадания в клетку замыкается в кольцо.

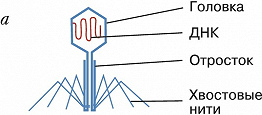
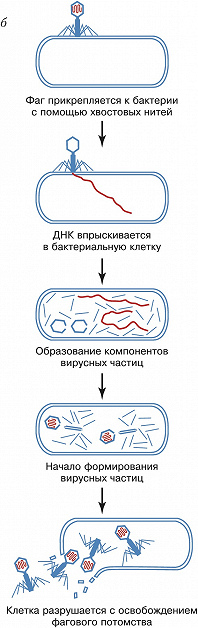
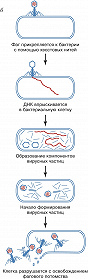
Б. прикрепляются к специфич. рецепторам на поверхности бактериальной клетки. Проникновение фагового генома в клетку сопровождается отделением нуклеиновой кислоты от большей части капсидных белков, которые остаются снаружи. С этого момента до появления первой зрелой фаговой частицы происходит строго упорядоченная во времени эскпрессия разных генов Б. – ранних и поздних. Группа ранних генов транскрибируется ферментом РНК-полимеразой: либо бактериальной, либо кодируемой фагом (в последнем случае её молекулы вводятся вместе с геномом); они контролируют репликацию генома, модификацию бактериальной мембраны. Поздние гены, как правило, транскрибируются с вновь образующихся геномов. При их участии появляются белки капсида, участвующие в сборке вириона, упаковке генетич. материала и обеспечивающие разрушение бактерии в определённое время. Образование отд. структурных элементов частиц Б. напоминает конвейерную сборку. На последнем этапе происходит упаковка генома в капсид.
В зависимости от характера взаимодействия с клеткой Б. делят на три группы. Заражение клетки вирулентными Б. (напр., T-чётными фагами кишечной палочки) приводит к разрушению (лизису) бактерии и высвобождению фагового потомства. Для умеренных Б. (в т. ч. фаги лямбда, Mu1, P1 и P2 энтеробактерий) возможны два пути развития – литический (сходный с развитием вирулентных фагов) и лизогенный (см. Лизогения), завершающийся установлением в клетке профага – скрытой неинфекционной формы вируса. При этом геном Б. встраивается в бактериальную хромосому или находится в свободном состоянии, так же как плазмида. Бактерия, несущая профаг, называется лизогенной; она устойчива к повторному заражению этим Б. При определённых условиях (облучение, действие химич. реагентов) может произойти активация профага и развитие Б. продолжится по литич. пути. Б. третьей группы (в т. ч. fd и M13) имеют непрерывный цикл развития. Они постоянно размножаются в клетке, а жизнеспособное фаговое потомство удаляется из неё через особые поры в клеточной оболочке. Однако не всегда удаётся однозначно отнести природные Б. к той или иной группе: умеренные фаги могут образовывать вирулентные мутанты, а геномы истинно вирулентных фагов (или их фрагменты) могут встраиваться в хромосомы бактерий.
Генетич. аппарат Б. подвержен изменениям, которые могут происходить в результате обмена отд. участками генома при совместном заражении одной и той же бактерии разными фагами, либо при участии мобильных генетических элементов. В геномах вирулентных Б. обнаружено множество генов, контролирующих синтез белков, сходных с белками самых разных организмов (в т. ч. патогенных бактерий, эукариот), а также разл. вирусов животных, что свидетельствует об участии Б. в миграциях генов в биосфере. Б. – важнейший фактор эволюции, приводящий к отбору фагоустойчивых вариантов бактерий, часто с изменёнными антигенными свойствами. Нередко в клетках обнаруживаются дефектные Б., которые возникают вследствие неправильного вырезания геномов умеренных Б. и/или при встраивании в хромосомы части генома Б. с участием мобильных генетич. элементов. Особое значение имеют Б., способные к трансдукции – переносу бактериальных генов в результате ошибочной упаковки в фаговую частицу фрагмента бактериальной ДНК (вместо или вместе с геномом Б.). Таким образом Б. обеспечивают быстрый обмен генами между разными штаммами (иногда видами) бактерий.
Б. широко используются в качестве модельных генетич. объектов благодаря работам М. Дельбрюка, А. Д. Херши и др., начавших в 40-х гг. 20 в. свои генетич. исследования и показавших применимость к Б. осн. положений классич. генетики. С помощью Б. было окончательно доказано, что генетич. материалом служит не белок, а ДНК, создана система анализа тонкой структуры генов, разработаны модели молекулярных механизмов мутагенеза. Благодаря Б. был окончательно подтверждён полуконсервативный характер репликации ДНК, впервые описан механизм интеграции вирусного генома в хромосому клетки-хозяина. Выяснение механизма рестрикции и модификации у Б. привело к открытию ферментов рестриктаз, что легло в основу генетич. инженерии. Б. используются для генетич. исследований бактерий, в качестве векторов для клонирования генов. Они нашли применение в медицине при лечении послеожоговых или раневых инфекций – фаготерапии, а также для идентификации патогенных бактерий.