КЛЕ́ТКА
-

Рубрика: Биология
-

-

Скопировать библиографическую ссылку:
КЛЕ́ТКА (лат. cellula, греч. ϰύτος), основная структурно-функциональная единица всех живых организмов. Представляет собой сложноорганизованную систему, состоящую из биополимеров (в т. ч. нуклеиновых кислот, белков) и др. соединений, способную к самоподдержанию и самовоспроизведению. К. может существовать как самостоятельный одноклеточный организм (бактерии, археи, простейшие, некоторые водоросли и грибы), так и в составе тканей и органов разл. многоклеточных организмов.
Историческая справка
Термин «К.» был предложен Р. Гуком (1665) для обозначения ячеек, которые он наблюдал с помощью увеличительных линз в срезах мёртвой пробковой ткани, представлявших собой лишь внешние оболочки растительных К. Позднее аналогичные «К.» Гука были обнаружены в составе разных частей растений и в тканях животных. Только в нач. 19 в. в связи с развитием микроскопии изменились общие представления о К.: главным в их организации стали считать не внешние оболочки («ячейки»), а содержимое – протоплазму (Я. Пуркине). В 1831 Р. Броун обнаружил и описал в протоплазме растит. К. постоянную структуру – ядро. На основании этих и многих др. наблюдений Т. Шванн сформулировал осн. положения клеточной теории (1839), одно из которых постулировало сходство строения К. растений и животных. Дальнейшее развитие клеточной теории связано с работами Р. Вирхова, доказавшего, что количество К. в организме увеличивается в результате клеточного деления (это правило было сформулировано им в его афористич. формуле – «Всякая клетка от клетки»). Создание клеточной теории явилось важнейшим событием в естествознании, одним из решающих доказательств единства всей живой природы. Основные её положения сохранили своё значение и в наше время. Совр. клеточная теория рассматривает К. как элементарную единицу живого; вне её нет жизни; увеличение числа К. происходит только путём деления исходных К.; многоклеточные организмы представляют собой сложные ансамбли К., объединённых в системе тканей и органов, и связаны межклеточными взаимодействиями; К. способны к метаболизму, использованию и трансформации энергии, чувствительности, изменчивости. Совокупность всех этих признаков можно обнаружить только на клеточном уровне.
Типы организации клеток
Среди живых организмов встречаются два типа строения К. — прокариотный и эукариотный. Прокариотные К. характерны для всех бактерий и архей; их размеры невелики (0,1–10 мкм); обычно это единичные, свободноживущие организмы, но некоторые из них могут образовывать колонии из одинаковых К. Они не имеют морфологически выраженного ядра – их геном (нуклеоид) в виде компактно упакованной кольцевой молекулы ДНК (т. н. бактериальной хромосомы) не отделён от цитоплазмы и структурно связан с плазматической мембраной, отграничивающей К.; снаружи прокариотная К. окружена клеточной стенкой (у архей т. н. S-слоями либо оболочкой, сходной с клеточной стенкой грамположительных бактерий); у некоторых прокариот клеточная стенка отсутствует. В цитоплазме (протоплазме) К. имеются вакуоли, многочисл. ферменты, обеспечивающие метаболизм, рибосомы, элементы цитоскелета; в К. могут находиться также и внехромосомные генетич. элементы – ДНК плазмид. Деление прокариотной К. — бинарное (путём образования перегородки), происходит после репликации ДНК.
Эукариотные К. свойственны всем остальным организмам. Они в 10–100 раз крупнее прокариотных; содержат морфологически выраженное ядро, отграниченное от цитоплазмы мембранной ядерной оболочкой и представляющее собой сложную систему, обеспечивающую хранение, воспроизведение и реализацию генетич. информации, содержащейся в линейных молекулах ДНК, заключённых в хромосомах. В цитоплазме, ограниченной плазматич. мембраной, выделяют жидкую плазму (гиалоплазма, или цитозоль), участвующую в промежуточном метаболизме К., а также множество спец. обязательных (наряду с ядром) структурных образований (органелл), каждая из которых выполняет определённые специфич. функции (сходные у разл. эукариотных К.). Рибосомы осуществляют синтез белка, митохондрии обеспечивают К. энергией, пластиды растительных К. (в т. ч. хлоропласты) участвуют в синтезе АТФ и фотосинтезе. Митохондрии и хлоропласты содержат собственный генетич. аппарат, что послужило основанием для возникновения гипотезы симбионтного происхождения эукариотных К. (см. Симбиогенез). Часть органелл, окружённых мембраной [эндоплазматическая сеть (эндоплазматич. ретикулум), аппарат Гольджи, эндо- и экзоцитозные вакуоли, лизосомы, пероксисомы], образуют вакуолярную систему. Кроме того, для К. эукариот характерно наличие сети белковых нитей, пронизывающих цитоплазму, — цитоскелет (опорно-двигательная система К.). У К. растений и грибов снаружи плазматич. мембраны расположена клеточная стенка, отсутствующая в К. животных. Обязательным компонентом цитоплазмы К. животных является клеточный центр, в состав которого входит центриоль. Соматические К. эукариот делятся путём митоза, сопровождающегося образованием спец. аппарата деления – клеточного веретена, с помощью которого происходит равномерное распределение хромосом (после их удвоения) строго по двум дочерним клеткам, в которых сохраняется исходный диплоидный набор хромосом. Продолжительность жизни К. от деления до деления составляет клеточный цикл. Половые К. (гаметы) у растений и животных формируются в ходе мейоза (в каждой К. оказывается гаплоидный набор хромосом).
Сходство и разнообразие клеток
Несмотря на мн. морфологич. и функциональные особенности, прокариотные и эукариотные К. во многом сходны, что обусловлено общностью структурно-молекулярной организации их внутриклеточных структур, которые образуют в К. некое сопряжённое единство и функционально взаимозависимы: и прокариотные, и эукариотные К. окружены плазматич. мембраной, выполняющей барьерную, транспортную и рецепторную функции; оба типа К. участвуют в процессах, связанных с поддержанием самой живой системы (напр., синтеза нуклеиновых кислот и белков, в биоэнергетических). Размножение К. происходит только путём деления исходной клетки, причём прокариотные К. используют аппарат деления К., напоминающий митотический.
Одновременно (при сходстве в строении внутриклеточных структур) эукариотные К. многоклеточных организмов могут различаться по форме, строению и размеру (даже в пределах одной особи, напр. К. эпителия, мышц, нервной системы, крови). Такое разнообразие К. можно объяснить специализацией функций, выполняемых разл. К., что является результатом избирательной активности разных генов в разных К. по мере развития многоклеточного организма (см. Дифференцировка). Показано, что любая К. такого организма обладает одинаковым полным набором генетич. материала, всеми возможностями для работы любого гена (т. е. полипотентна), но в разных К. одни и те же гены могут находиться в активном или в репрессивном состоянии. Всегда активны гены, определяющие синтез белков и нуклеиновых кислот, необходимых для поддержания жизни самой К., но в К. разных тканей, кроме того, активны специфич. гены, определяющие свойства и осн. клеточные функции, характерные именно для К. данной ткани. Регуляция же активности спец. генов определяется межклеточными (см. Межклеточные взаимодействия), гормональными, нервными (для животных) формами химич. регуляции.
Т. о., в многоклеточном организме К. является единицей развития, единицей функционирования в разл. органах и тканях и первичным объектом патологич. изменений жизнедеятельности. У эукариот развитие организмов начинается с родоначальной К. (зиготы), потомки которой образуют сложные многоклеточные ансамбли тканей и органов. Любые функции многоклеточного организма – результат работы отд. К. в разных органах. Нарушение целостности К. или изменение их свойств – осн. причина всех без исключения заболеваний. Естественное разрушение клеток происходит в ходе реализации генетически детерминированной программы – апоптоза.
Наука о К. — клеточная биология. Для изучения К. используют разл. виды микроскопии, культивирование К. (см. Культура клеток и тканей), разл. методы клеточной инженерии, а также многочисл. методы биохимии, биофизики, молекулярной биологии.
-
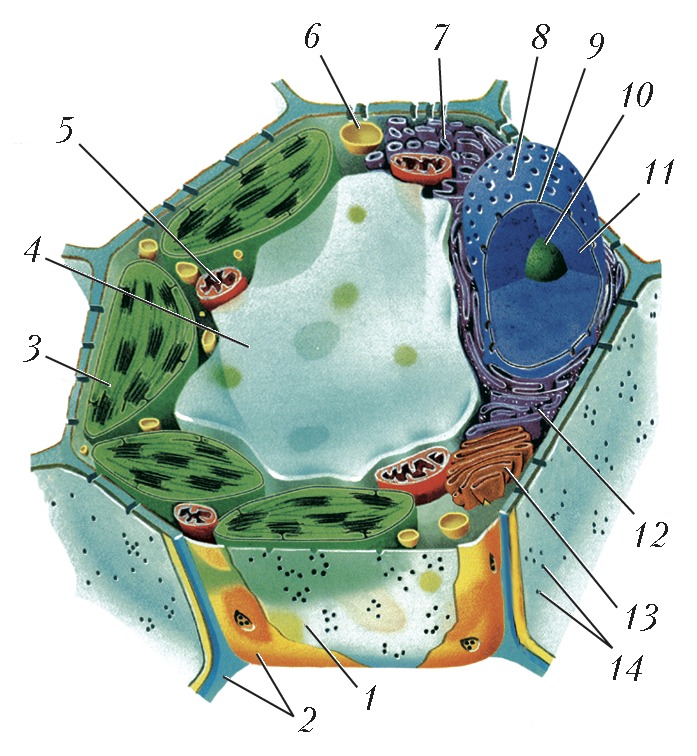
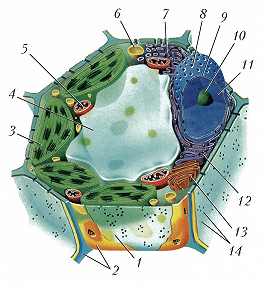
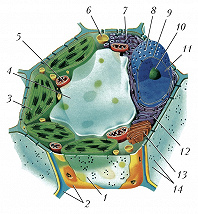 Схема строения растительной клетки: 1 – плазматическая мембрана; 2 – клеточная стенка; 3 – хлоропласты; 4 – вакуоль; 5 – митохондрия; 6 – лизосома; 7 – гладкий эндоплазматический ретикулум; 8 – ядерная пора; 9 – ядерная мембрана; 10 – ядрышко; 11 – ядро; 12 – гранулярный эндоплазматический ретикулум; 13 – аппарат Гольджи; 14 – плазмодесма.
Схема строения растительной клетки: 1 – плазматическая мембрана; 2 – клеточная стенка; 3 – хлоропласты; 4 – вакуоль; 5 – митохондрия; 6 – лизосома; 7 – гладкий эндоплазматический ретикулум; 8 – ядерная пора; 9 – ядерная мембрана; 10 – ядрышко; 11 – ядро; 12 – гранулярный эндоплазматический ретикулум; 13 – аппарат Гольджи; 14 – плазмодесма. -
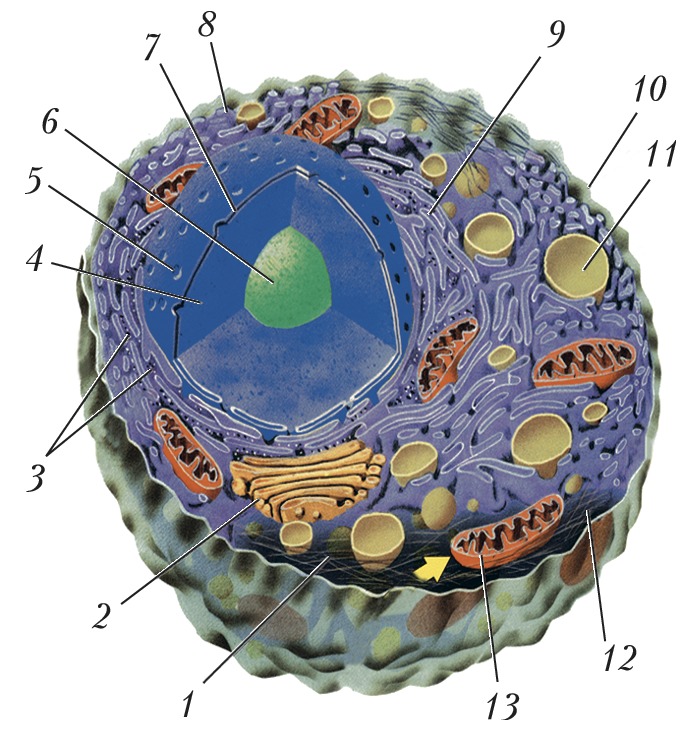
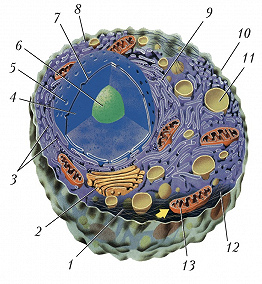
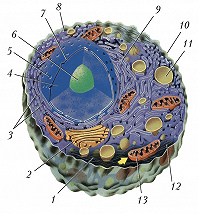 Схема строения животной клетки: 1 – микротрубочки; 2 – аппарат Гольджи; 3 – рибосомы; 4 – ядро; 5 – ядерная пора; 6 – ядрышко; 7 – ядерная мембрана; 8 – гладкий эндоплазматический ретикулум; 9 – гранулярный эндоплазматический ретикулум; 10 – плазматическая мембрана; 11 – лизосома; 12 – цитоплазма; 13 – митохондрия.
Схема строения животной клетки: 1 – микротрубочки; 2 – аппарат Гольджи; 3 – рибосомы; 4 – ядро; 5 – ядерная пора; 6 – ядрышко; 7 – ядерная мембрана; 8 – гладкий эндоплазматический ретикулум; 9 – гранулярный эндоплазматический ретикулум; 10 – плазматическая мембрана; 11 – лизосома; 12 – цитоплазма; 13 – митохондрия. -
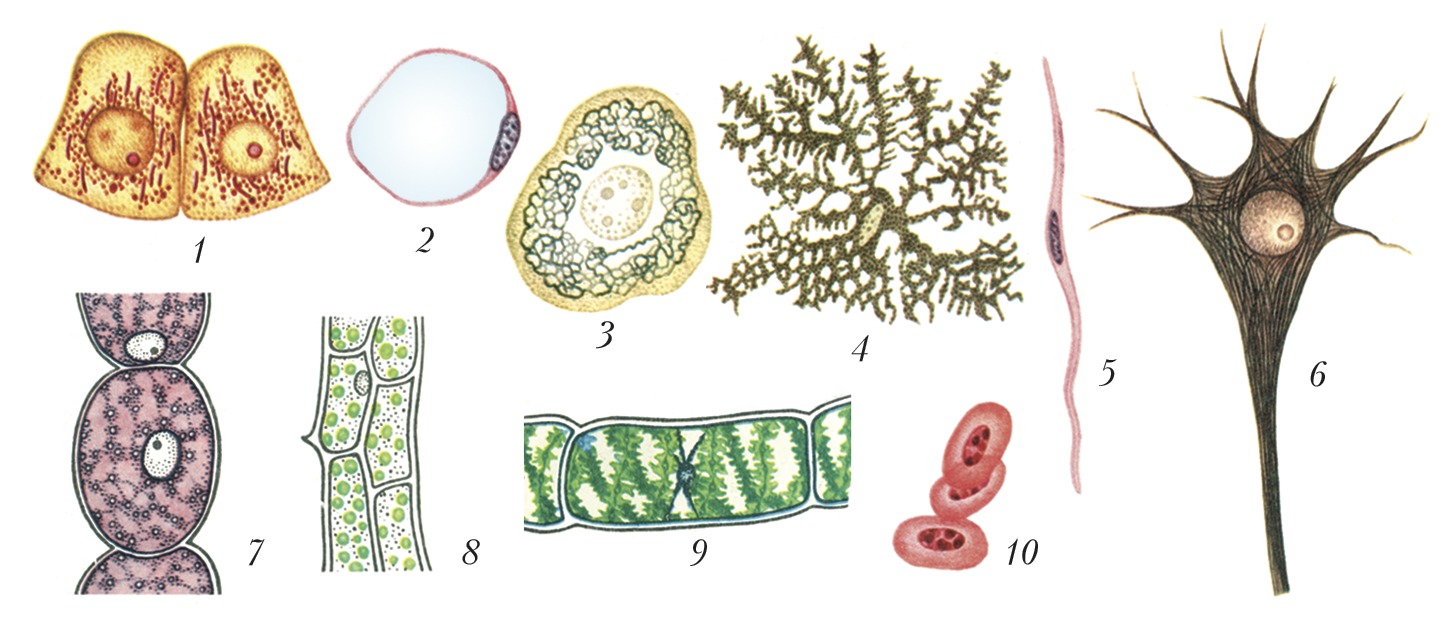
 Разнообразие животных и растительных клеток (клетки окрашены разными способами, поэтому видны не все детали): 1 – клетки почки лягушки, видны удлинённые митохондрии и ядро; 2 – жировая клетка из подкожной клетчатки крысы с прижатым к плазматической мембране ядром и крупной жировой каплей; 3 – чувствительная клетка спинномозгового ганглия человека с ядром и аппаратом Гольджи; 4 – пигментная клетка (хроматофор) аксолотля, заполненная гранулами пигмента; 5 – клетка гладких мышц кишечника человека; 6 – клетка коры головного мозга человека – нейрон с длинным отростком (аксоном) и ветвящимися короткими (дендритами), видны ядро с ядрышком и нитчатые структуры (нейрофибриллы) в цитоплазме; 7 – клетка волоска тычиночной нити традесканции; 8 – клетки листа элодеи; 9 – клетка водоросли спирогиры; 10 – ядерный эритроцит курицы.
Разнообразие животных и растительных клеток (клетки окрашены разными способами, поэтому видны не все детали): 1 – клетки почки лягушки, видны удлинённые митохондрии и ядро; 2 – жировая клетка из подкожной клетчатки крысы с прижатым к плазматической мембране ядром и крупной жировой каплей; 3 – чувствительная клетка спинномозгового ганглия человека с ядром и аппаратом Гольджи; 4 – пигментная клетка (хроматофор) аксолотля, заполненная гранулами пигмента; 5 – клетка гладких мышц кишечника человека; 6 – клетка коры головного мозга человека – нейрон с длинным отростком (аксоном) и ветвящимися короткими (дендритами), видны ядро с ядрышком и нитчатые структуры (нейрофибриллы) в цитоплазме; 7 – клетка волоска тычиночной нити традесканции; 8 – клетки листа элодеи; 9 – клетка водоросли спирогиры; 10 – ядерный эритроцит курицы.