ФОТОСИ́НТЕЗ
-

Рубрика: Биология
-

-

Скопировать библиографическую ссылку:
ФОТОСИ́НТЕЗ (от фото... и синтез), преобразование зелёными растениями и некоторыми бактериями энергии света в энергию химич. связей органич. веществ. Поглощение света происходит при участии хлорофиллов (бактериохлорофиллов) и некоторых др. вспомогательных фотосинтетич. пигментов (каротиноиды, фикобилины), присутствующих в хлоропластах и хроматофорах клеток. Исключение составляют галобактерии, для которых характерен Ф. с помощью мембранного белка бактериородопсина. У зелёных растений, водорослей и цианобактерий исходными неорганич. соединениями, необходимыми для синтеза органич. вещества, являются CO2 и H2O. В основе Ф. лежат окислит.-восстановит. реакции, в ходе которых электроны переносятся от донора-восстановителя (вода, водород и др.) к акцептору (СО2, ацетатион) с образованием восстановленных соединений (углеводы) и выделением молекулярного кислорода (O2). Фотосинтезирующие бактерии, не использующие в качестве донора H2O, кислород не выделяют (см. Хемосинтез).
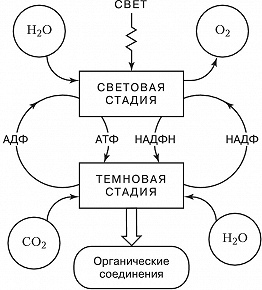
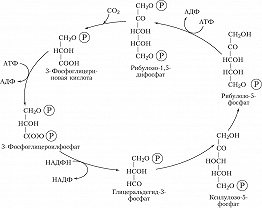
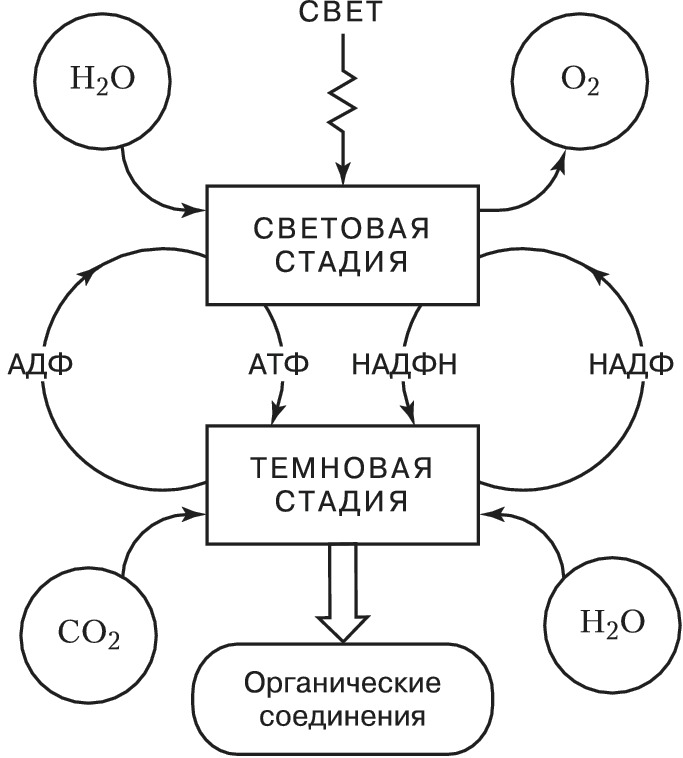
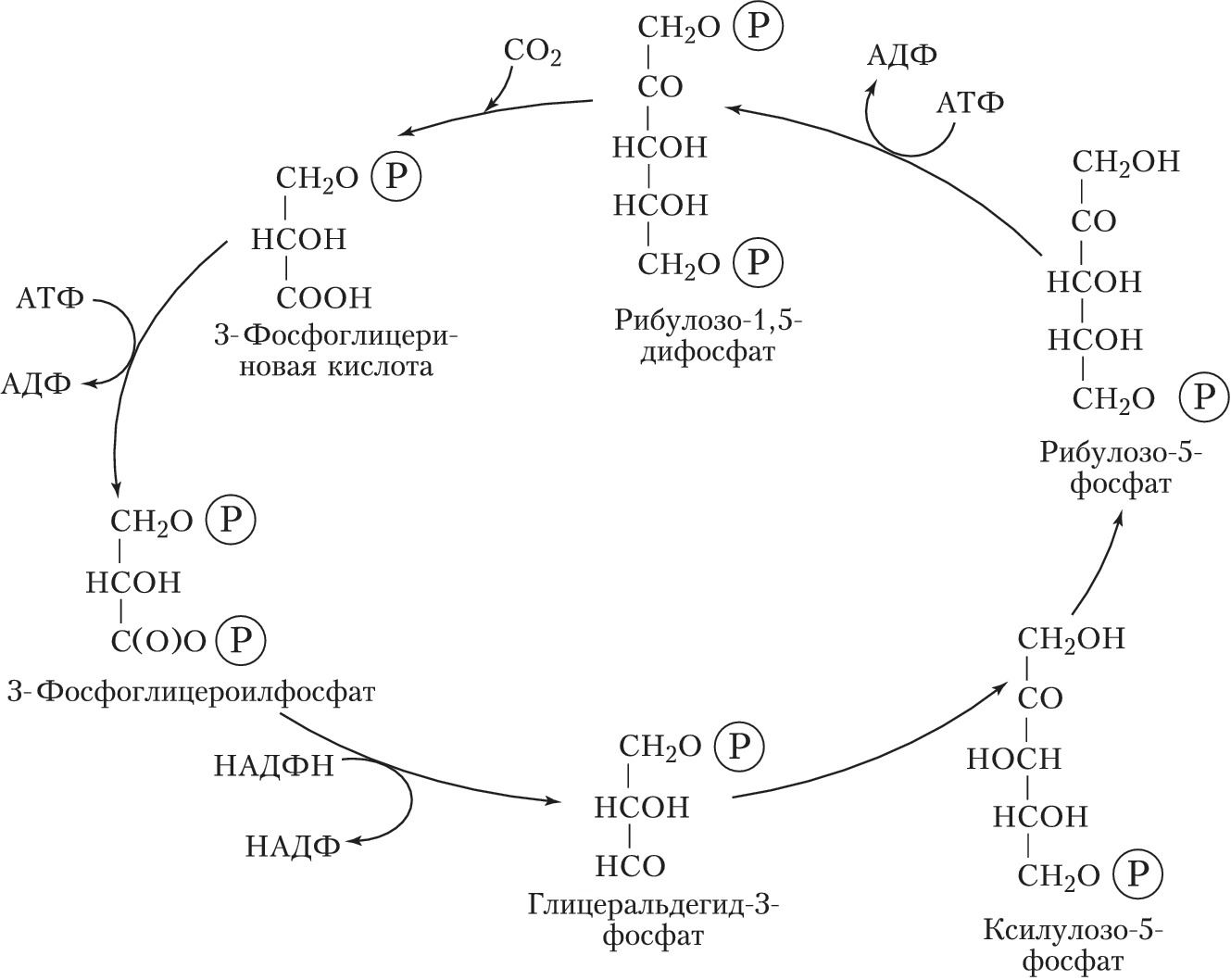
Ф. высших растений, водорослей и цианобактерий делят на две стадии – световую и темновую (рис. 1). Первая стадия (световые реакции) требует энергии света и осуществляется в мембранных структурах хлоропластов – тилакоидах, в которые встроены молекулы пигментов и переносчиков электронов – пластохинона и пластоцианина. Энергия света, поглощённая пигментами, поднимает их электроны на более высокий энергетич. уровень: они приобретают избыток энергии (энергию возбуждения). Следствием этой стадии является распад молекулы воды (фотолиз) на водород и кислород и восстановление никотинамидадениндинуклеотидфосфата (НАДФ+) – кофермента дегидрогеназ – до НАДФН. Одновременно с этим благодаря формированию на мембранах тилакоидов электрохимич. градиента протонов происходит образование из АДФ богатого энергией АТФ (фотофосфорилирование); O2 выделяется в атмосферу. Темновая стадия (темновые реакции) протекает в строме хлоропластов. Она активируется светом и может протекать на свету, поэтому только условно может быть названа темновой. В темновых реакциях для восстановления CO2, поступающего через межклетники листа в хлоропласты, используется энергия, запасённая в НАДФН и АТФ световых реакций. Фиксация CO2 и образование углеводов происходит в ходе последовательных ферментативных превращений – цикла Калвина (назван по имени описавшего его М. Калвина). Первая реакция цикла (см. рис. 2) – при участии фермента рибулозодифосфаткарбоксилазы CO2 присоединяется к молекулам рибулозодифосфата (содержат 5 атомов углерода, С5); из образовавшихся шестиуглеродных соединений получаются молекулы 3-фосфоглицериновой кислоты (С3). Она восстанавливается (при участии АТФ и НАДФН) до триозофосфатов, из которых образуется конечный продукт – глюкоза. Вместе с тем часть триозофосфатов претерпевает процесс конденсации и перестроек, превращаясь в рибулозомонофосфат, который с участием АТФ фосфорилируется до рибулозодифосфата – первичного акцептора CO2, что и обеспечивает непрерывную работу цикла. Растения, Ф. которых происходит через образование трёхуглеродных соединений, получили назв. С3-растений. В некоторых растениях (кукуруза, сахарный тростник и др.) первоначальное превращение углерода идёт не через трёхуглеродные, а через четырёхуглеродные соединения (С4-растения). Акцептором СО2 в клетках мезофилла таких растений служит фосфоенолпируват. Продукты его карбоксилирования – яблочная или аспарагиновая кислоты – диффундируют в обкладочные клетки сосудистых пучков, где декарбоксилируются с освобождением СО2, который и поступает в цикл Калвина. Преимущества такого «кооперативного» метаболизма обусловлены тем, что фосфоенолпируваткарбоксилаза при низкой концентрации СО2 более активна, чем рибулозодифосфаткарбоксилаза, и, кроме того, в обкладочных клетках с пониженной концентрацией O2 слабее выражено окисление рибулозофосфата и сопутствующая ему потеря энергии (до 50%). Поэтому С4-растения характеризуются высокой фотосинтетич. продуктивностью.
Ф. – единственный процесс в биосфере, ведущий к увеличению свободной энергии биосферы за счёт внешнего источника – Солнца и обеспечивающий существование как растений, так и всех гетеротрофных организмов, в т. ч. человека. Ежегодно в результате Ф. на Земле образуется 150 млрд. т органич. вещества и выделяется ок. 200 млрд. т свободного О2. Кругооборот О2, углерода и др. элементов, вовлекаемых в Ф., создал и поддерживает совр. состав атмосферы. Ф. препятствует увеличению концентрации СО2 в атмосфере, предотвращая перегрев Земли вследствие парникового эффекта. Кислородно-озоновый экран атмосферы необходим для защиты от УФ-излучения. Запасённая в продуктах Ф. энергия (в виде разл. видов топлива) является осн. источником энергии для человека. Предполагается, что в энергетике будущего Ф. может занять одно из первых мест в качестве неиссякаемого и незагрязняющего среду источника энергии (создание «плантаций» быстрорастущих растений с последующим использованием растит. массы для получения тепловой энергии или переработки в высококачественное топливо). Не менее важна роль Ф. как основы получения продовольствия, кормов, технич. сырья. Несмотря на высокую эффективность начальных стадий (ок. 95%), в биомассу переходит лишь менее 1–2% солнечной энергии; потери обусловлены неполным поглощением света, лимитированием процесса на биохимич. и физиологич. уровнях. Обеспечение растений водой, минер. питанием, СО2, селекция сортов с высокой эффективностью Ф., создание благоприятной для светопоглощения структуры посевов и др. пути используются в целях реализации значит. резервов фотосинтетич. продуктивности. Для ряда культур оправдано выращивание при полном или частичном искусств. освещении, биотехнологич. способы получения растит. массы (особенно одноклеточных организмов), аквакультура для некоторых водорослей и т. д. В связи с этим особенно актуальными становятся разработка теоретич. основ управления Ф., исследование Ф. как целостного процесса, закономерностей его регулирования и адаптации к внешним условиям.